СЕЛЕКЦИЯ И ВОСПРОИЗВОДСТВО
Еськов ЕЖ.
Российский государственный аграрный заочный университет
В настоящее время семейство пчелиных объединяет 520 родов. Наибольшим их таксономическим разнообразием отличаются Неотропики, Нео- и Палеарктика, в которых известно обитание 315, 260 и 243 родов соответственно. Самое низкое представительство пчелиных в Австралийской области (описано всего 18 родов). Род Apis включает шесть видов, среди которых наибольший ареал занимает медоносная пчела (Apis mellifera). Предковая форма рода Apis не установлена. Возможно, он сформировался среди вымерших в верхнем эоцене пчел рода Electrapis. Эти пчелы, среди известных ископаемых форм, по комплексу морфологических признаков имеют сходство с представителями рода Apis, но по признакам жилкования они близки к шмелям (Bombus).
Остатки двух видов пчел, сходных с современными видами А. dorsata и A. laboriosa, обитающими в тропиках пчел, обнаружены в отложениях, возраст которых составляет 6-10 млн. лет. Что же касается места возникновения (прородины) Apis mellifica, то оно пока не определена. Этот вид пчел мог сформироваться в Юго-Восточной Азии. Однако, не исключена возможность его евроафриканского происхождения.
Современный род Apis дифференцируется на три подрода: 1) Apis — A. mellifera и А. сегапа; 2) Megapis — A. dorsata и A. laboriosa; 3) Micrapis — A. florea и A. andreniformis. Очевидно, образование видов в каждой из трех филетических ветвей рода Apis (Apis, Megapis и Micrapis) происходило по принципу дивергенции. Однако в дальнейшем они развивались по принципу парафилии. Этому способствовало сходство наследственной основы и совпадение направлений отбора в процессе адаптации к сходным условиям, что повлияло на сближение морфофизиологических признаков.
Среди пчел рода Apis, к которому принадлежит медоносная пчела, по эвритермности (способности жить в широком диапазоне температур) выделяется медоносная пчела. Ее ареал распространяется от тропиков до Полярного круга. Исходно, медоносная пчела заселяла Старый свет (Европу, Африку и Азию) и только в середине второго тысячелетия стала распространяться в Новый Свет. Это связано с высокими адаптивными свойствами вида. Немаловажное значение в расселении пчел имеет также антропическая деятельность.
Первые обобщения знаний о поведении пчел содержатся в биологических трактатах Аристотеля, написанных в 4 в. до нашей эры. Однако долгое время после этого основные принципы организации пчелиной семьи оставались нераскрытыми. Значительные достижения в изучении биологии пчел были выполнены на границе XVI-XVII вв. В конце XVI века Л.М. де Тореесу удалось обнаружить что «царица» пчелиной семьи выполняет репродуктивную функцию. В начале XVII века Г. Батлер определил половую принадлежность трутней, а Р. Рамнат установил пол рабочих пчел. Бурное изучение биологии пчел происходило в XX веке и продолжается в настоящее время.
Медоносная пчела была и остается предметом пристального изучения этологов, физиологов, энтомологов и специалистов аграрного комплекса. Достижения последних десятилетий в области изучения этологии и физиологии органов чувств пчелы повлияли на успешное развитие биологии. Расшифровка уникальных принципов поддержания гомеостаза семьи, механизмов пространственной ориентации и сигнализации пчел открыла новые перспективы для управления их поведением. Особого внимания заслуживают сведения о терморегуляции и системе внутригнездовых коммуникаций.
Механизм терморегуляции основан у пчел на использовании мобильного этолого-физиологического комплекса, включающего изменение тепловыделения, теплоизоляции или охлаждения. Использование того или иного средства терморегуляции зависит от экологической ситуации. На охлаждение пчелы реагируют активизацией термогенеза и усилением теплоизоляции посредством агрегирования вокруг зон, занятых расплодом. Связанное с этим уплотне¬ние пчел и уменьшение занимаемого ими внутригнездового объема способствует снижению тепловых потерь за счет уменьшения излучения и теплообмена. Эффективным средством охлаждения служит аэрация гнезда (пчелы-вентилировщицы машут крыльями, создавая направленный поток воздуха) и связанная с этим интенсификация испарения воды через покровы их тела. Частота взмахов крыльями возрастает с повышением температуры. Соответственно этому возрастает поток воздуха, охлаждающего гнездо. Так, при 35-35.5оС пчелы машут крыльями с частотой 75-85 Гц, а при 41-42оС она воз¬растает до 145-155 Гц.
Активная роль в охлаждении гнезда при высокой внешней тем¬пературе принадлежит пчелам, специализирующемся временно на доставке воды (пчелы-водоносы). В другое время эти пчелы могут заниматься доставкой нектара, пыльцы или смолистых выделений растений.
У пчел, в отличие от других видов высокоорганизованных социальных насекомых (термитов и муравьев) отсутствует строгая функциональная дифференциация взрослых особей. Однако в период активной жизни среди рабочих особей прослеживается более-менее выраженная функциональная изменчивость, связанная с возрастом. Обычно в кормлении расплода участвуют пчелы младших возрастных групп, а доставке корма и воды — старших. В зависимости от биологической ситуации пчелы-фуражиры могут становиться водоносами, сторожами, кормилицами, участвовать в строительстве сот и других внутри- и внегнездовых работах.
Поведение и продуктивность полетов пчел-водоносов зависит от температуры. Минимальная температура, при которой пчелы могут вылетать к известным им источникам воды, находящимся на небольшом расстоянии (нескольких метров от улья), составляет 6оС, что нередко происходит в периоды длительных весене-летних похолоданий. Эти пчелы перед вылетом берут в среднем 7 мг меда (макси¬мально — 40 мг).
В течение периода зимовки (при отсутствии расплода) стабильность внутригнездовой температуры понижается (Еськов, Тобоев, 2009,2010). В генерации тепла зимующими пчелами доминирующая роль принадлежит той их части, которая локализуется в тепловом центре. Вокруг него агрегируются пчелы, выполняющие преимущественно теплоизолирующую функцию. Плотность их агрегирования повышается, а локомоторная активность понижается соответственно удалению от теплового центра и усилению охлаждения. На периферии (за исключением зоны над тепловым центром) гиподинамия усиливается настолько, что может приближаться, а у некоторой части пчел достигать состояния холодового оцепенения. Эти пчелы отогреваются и активизируются при наслоении на них разогретых пчел, покидающих тепловой центр. Миграция пчел в теплую часть гнезда, а из нее на периферию происходит в течение всей зимовки, чем порождается высокая вариабельность их локомоторной и трофической активности.
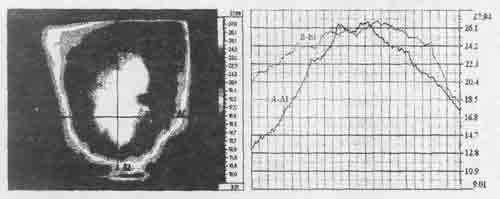
Рис.1. Распределение тепловых полей в центральном межсотовом скоплении пчел при -2°С: А — термограмма, Б — температурные профили в сечениях А-А1 и В-В1.
Внутригнездовые сигналы связи. Пчелы используют сложный комплекс динамических (двигательных), химических и акустических сигналов связи. В начале XX в была обнаружена связь между формой стереотипных движений (танцев), совершаемых пчелами на сотах после обнаружения обильного источника корма (Еськов, 1979). Траектория движений танцовщиц, длительность всего танцевального цикла и его «виляющей» фазы (периода, в течение которого пчела машет брюшком в дорсальной плоскости), зависят от расстояния до цели полета. По направлению «виляющей» фазы определяется направление от улья к источнику корма.
Обычно пчела совершает подряд несколько циклов танцевальных движений. В круговом танце она передвигается по окружности и, как правило, не закончив виток, резко разворачивается и продолжает движение в противоположном направлении. С этого момента начинается следующий цикл танца. В начальной фазе восьмерочного танца пчела движется по прямой линии, совершая маятникообраз-ные движения брюшком. Завершив пробег, она возвращается в исходное или близкое к нему положение по дугообразной траектории. Направление движения по ней перед очередным выходом на прямолинейный пробег большей частью меняется на противоположное.
Впервые на специфические динамические позы у пчел обратил внимание М. Спинцер (Spitzner) и опубликовал свои наблюдения в 1788 г. Позже этим занимался К. Фриш и его последователи, посвятившие изучению танцев пчел множество публикаций. Но эти исследования не давали убедительного объяснения биологической роли пчелиных танцев. Несмотря на то, что продолжительность виляющей фазы коррелирут с расстоянием до цели полета, а направление к ней определяет вектор перемещения пчелы в то время, когда она ма¬шет брюшком, визуальное использование всех этих динамических поз в пчелином гнезде невозможно по многим причинам. Прежде всего, потому, что в улье темно, а диапазон светового восприятия пчел сдвинут в область ультрафиолета. Контролировать движения танцовщиц со стороны невозможно из-за высокой плотности пчел на сотах. Поэтому сами по себе танцевальные движения пчел не могут нести информационной нагрузки. Она содержится в звуковых сигналах и электрических колебаниях, генерируемых статически заряженным брюшком танцовщицы.
Звуковой сигнал, используемый пчелами для указания расстояния до цели полета, представляет собой упорядоченный по амплитуде и частоте следования пакет импульсов, каждый из которых состоит в среднем из четырех звуковых периодов и длится около 15 мс. Паузы между импульсами имеют примерно такую же длительность. Импульсы следуют с частотой около 33 Гц. Длительность импульсов, как и пауз между ними, не связана с расстоянием до цели полета. С ним коррелирует длительность всего сигнала и количество составляющих его импульсов.
Наличие связи между длительностью звукового сигнала танцовщицы и расстоянием до цели полета указывает на способность пчел измерять длину преодолеваемого ими пути. Это доказано опытами, в которых у пчел-сигнальщиц, посещавших кормушку на определенном расстоянии от улья, укорачивали на 15 — 18% длину левого переднего крыла. В результате непосредственно после ампутации части крыла длительность З.с.д., посещавших кормушку на расстоянии 200 м от улья, возрастала в среднем от 609±11 мс до 906±16 мс, а количество импульсов — от 18±0.5 до 21.1 ±0.5 мс. В дальнейшем пчелы адаптировались к изменению длины крыла, что выражалось в уменьшении наполнения зобиков примерно на 20%, а длительность сигнала стабилизировалась на уровне 864±19 мс.
Длительность звуковых сигналов и количество импульсов в нем является врожденным расовым признаком пчел. В частности у итальянских пчел, сигнализирующих об источнике корма, находящемся на расстоянии 200 м от улья составляет 635±13 мс. В течение этого времени пчела генерирует 21.2±0.4 импульса. Краинские пчелы в тех же условиях генерируют 12.9±0.3 импульса в течение 434±19 мс. Эти свойства звуковых сигналов наследственно запрограммированы и не изменяются в течении жизни пчел одних рас в семьях других рас.

Рис, 2. Временная структура сигнала итальянских (А) и краинских (Б) пчел, посещающих источники корма на расстоянии 30 (а), 100 (б) и 200 мот улья (В)
Для восприятия низкочастотных сигналов у пчел получили развитие быстроадаптирующиеся трихоидные сенсиллы. Они обеспечивают ближнюю связь, что необходимо для быстрого взаимодействия в густонаселенном пчелином жилище. Повышению надежности внутригнездовой коммуникации способствует восприятие трихоидными сенсиллами низкочастотных электрических колебаний. Они генерируются вместе со звуками летальным аппаратом и другими локомоторными органами.
Заключение.
Развитие большого комплекса адаптации в филогенезе медоносной пчелы сопряжено с эволюцией социальности. Она возникла в процессе прогрессирующего усиления взаимосвязи и взаимозависимости самок-основательниц с их потомством. Консолидации семьи сопутствовала утрата индивидуальных преимуществ и конкуренции между ее членами. Отбор благоприятствовал сохранению и селективному размножению признаков, способствовавших повышению адаптивности всей семьи. На этом пути происходило превращение пчелиной семьи в элементарную единицу естественного отбора.
Литература:
1. Еськов Е.К. Акустическая сигнализация общественных насекомых. — М.: Наука, 1979. — 209 с.
2. Еськов Е.К., Тобоев В.А. Экзо- и эндогеннозависимые флуктуации терморегуляторной активности медоносной пчелы// Известия РАН. Сер. биологическая. — 2009. — №2. — С. 249-256.
3. Еськов Е.К., Тобоев В.А. Генерация, накопление и рассеяние тепла в скоплениях агрегирующихся насекомых// Журн. общей биологии. — 2009. — Т. 70. — № 2. — С. 110-120.
4. Еськов Е.К., Тобоев В.А. Анализ статистически однородных фрагментов акустических шумов, генерируемых скоплениями насекомых// Биофизика. -2010. -Т.55. -№1. -С. 113-125.
5. Расницын А.П. Происхождение и эволюция перепончатокрылых насекомых. М.: Наука, 1980. — 191 с.